Kommentar til boka 'Evolusjon eller Kristen tro' -ja takk begge deler
Kap. 3 Argument I om ID. Arg. 4: Evolusjon forklarer mye, men kommer til en grense den ikke kan passere
Utsagn i boka: "Fordelen med så klar tale (som Behes) er at den kan testes."

Kommentar: Endelig en innrømmelse om at ID-påstander kan testes. Og det er mulig generelt, ikke bare her: For å motbevise ID, er det tilstrekkelig å peke på én mekanisme i naturen som naturlig ('blindt') har utviklet kompleks, spesifisert informasjon. For å motbevise ET, må en vise at det ikke finnes noen måte livet kan oppstå/utvikle seg på, noe som er en bortimot uoverkommelig oppgave. For øvrig er det igjen ID som må lide under slike demarkasjonskriterier. Noen disipliner f.eks. innen teoretisk fysikk, som strengteori, er ikke testbare i det hele. Men i forbindelse med testbarhet, ytres en tvil om hvorvidt 'det samme kan sies om tusen år'? Men det ligger i vitenskapens vesen at ting kan endre seg, og her uttaler Behe seg ut fra historiske erfaringer, verden over, i forbindelse med malaria-epidemi.
Påstand i boka: "Blant det Behe bommer på her, er at evolusjonen ikke kan grantere at alle grupper vil utvikle nye funksjoner på kort tid, til tross for at den her nettopp har sørget for en måte å bekjempe malaria på. Videre er mangelen på slike mutasjoner i én gruppe ikke bevis på at slikt ikke kan skje i noen grupper."
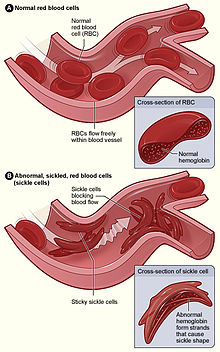
Kommentar: Om man etterprøver Davidsens syn på at evolusjonen 'har sørget for en måte å bekjempe malaria på', kan man finne ut dette: Sigdcelle-anemi er ett av de mest kjente eksemplene på 'positive' mutasjoner som opptrer i enkelt dose. Men i dobbel dose må denne mutasjonen operere på egne premisser, og da først røper den hva som bor i den. Sigdcelle-anemi i dobbel dose får man, selvsagt, om begge foreldre er bærer av mutasjonen. Det kommer da for en dag at mutasjonen kan volde komplikasjoner i form av blokkering av trange blodkar, med økt risiko for dødelighet. Dette gir et klart bevis på at sigdcelle-mutasjonen er en ugunstig mutasjon -lenke.
Bilde 1og 2. Sigdcelleanemi er en arvelig sykdom, som kan føre til blokkering av trange blodkar.
Som Davidsen skriver "evolusjon hemmes kraftig når det trengs to mutasjoner for å få en adaptiv fordel. Er ikke den første mutasjonen nyttig, spres den for lite i populasjonen til at noen kan få den andre nødvendige mutasjonen tilfeldig, mens de fortsatt har den første. Dermed må begge mutasjonene skje samtidig i samme genom, noe som skjer ytterst sjelden."
Våre proteiner er delikate mekanismer, som tåler lite slingringsmonn. Hemoglobin, som transporterer røde blodcellers oksygen rundt i kroppen, har aldri blitt utkonkurrert av noe funnet opp i et laboratorium. Om hemoglobin-molekyl ødelegges i mutasjoner, kan det føre til sigdcelle-anemi, som altså i dobbel dose er dødelig. Det samme er tilfellet for de fleste proteiner. De fleste proteiner er så gode som de kommer til å bli, blir det faktisk nå hevdet av flere evolusjonister.
F.eks. er Dan Tawfik, Weizmann Institute, en talsmann for denne idéen. Han mener at 'bred-spesifikke' enzymer tjente som 'originator' for dagens spesialiserte enzymer. (1) Til sin fordel gjenkjenner Dan Tawfik vansken med å forklare hvordan de formodede 'bred-spesifikke' enzymer oppsto i første omgang. Det måtte også ha vært skikkelige enzymer, foldede proteiner, med geometrisk komplekse aktive lokasjoner. Det er dermed uklart hva som oppnås ved å foreslå dem som forløpere. Tawfiks egen diagnose om dette, er overraskende ærlig. I følge ham har evolusjonsteorien dette 'Catch-22': Ingenting utvikler seg uten at det allerede eksisterer' (2). En kan altså ikke forvente et virksomt Y ut av noen prosess, om ikke et virksomt X gikk inn i den.
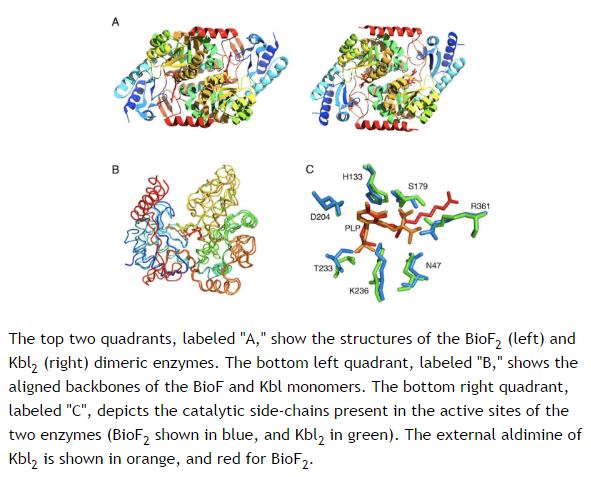
Bilde 2. To rimelige like proteiner med ulik funksjon
Noe som må synes som ett tankekors for evolusjonister, er at natrulig seleksjon påstås å ha ført til enorme forandringer i form av makro-evolusjon i en fjern fortid, mens når det testes på et laboratorium i våre dager, så kan ikke mutasjons-framkalte endringer få to nesten identisk like proteiner (Kbl og BioF) til å oppnå lik funksjon (3,4). Mange vitenskapsfolk mener faktisk at de to tingene kan kombineres. Det nåværende svaret fra disse evolusjonistene, er at naturlig seleksjon er 'offer for sin egen suksess'. Det innebærer at naturlig seleksjon nå oppfattes som så god til å tilpasse organismer til sine omgivelser, at de nådde sluttpunkter: Organismer som er så gode til det de er, at de ikke lenger undergår evolusjonære endringer.
(I stykket under benyttes forkortelsen NDT for Neo-Darwinistisk Teori.)
Påstand i boka: "Men det hele avhenger av at viktige evolusjonære steg faktisk krever flere samtidige mutasjoner, noe Behe ikke klarer å vise må være tilfellet. Ikke minst antyder klorkinineksempelet at dette faktisk er mulig."
Svarforslag: For å ta det siste først, la oss se på hva som skjer i forbindelse med klorkinineksempelet:
Den enkleste måten å endre et protein er ved punktmutasjon, der én aminosyre blir erstattet med en annen, i en posisjon i proteinet. Det er 20 ulike aminosyrer i proteiner. Det innebærer at om bare 5-6 posisjoner endret til de rette, de som tillot proteinet å binde, så ville det være en begivenhet som kunne tilsvare påstått virkelighet. 20 multiplisert med seg selv 5 ganger gir 3 millioner, og 20 multiplisert med seg selv 6 ganger gir 60 millioner. 5-6 mutasjoner synes ikke særlig mye, når proteiner består av hundrevis av aminosyrer, men det er et viktig poeng at det kreves 5-6 samsvarende (kontinuerlige') mutasjoner, bare for å få to proteiner til å binde seg til hverandre. Ett manglende steg viser seg å være mye vanskeligere når stegene skal være kontinuerlige. Om multiple steg mangler, blir vanskegraden økt eksponentielt. Så kan vi i raushetens navn si at omtrent en tredel endringer i aminosyrer, er nøytrale. Da gjenstår 3-4 samsvarende aminosyre-endringer, som kan volde problemer om de skjer hver for seg. Tre eller fire samtidige aminosyre-endringer er som å droppe flere trappetrinn i en evolusjonær trapp. Det er 1-2 flere aminosyre-endringer enn det som trengtes for å oppnå klorokin- resistens -med kallenavn CCC (chloroquine-complexity-cluster) i malaria, noe som har reell sannsynlighet ved fødsel. M.a.o. blir det å generere én ny cellulær protein-protein binding minst av samme vanskegrad som utvikling av klorokin-resistens i malaria-parasitten.
Anta nå at for å oppnå en fordelaktig egenskap, så måtte en ha ikke bare én, men to nye protein-bindinger. Sannsynligheten for CCC er 1: (10 opphøyd i 20), der siste tall tilsvarer hundre milliarder, milliarder organismer. Det er flere enn antall pattedyr som har eksistert på jorda. Under ellers like forhold, ville sannsynligheten for å oppnå en dobbel CCC, være kvadratet av én CCC, eller 1: (10 opphøyd i 40). Det siste tallet er et høyere tall enn antall celler det sannsynligvis noensinne har eksistert på jorda. Statistikk dreier seg mye om gjennomsnitt, så en kan ikke hevde at det er logisk umulig, men det ville ikke være 'naturlig' å forvente to slike protein-bindinger etter neo-darwinistiske mekanismer, i løpet av jordas historie. På kortform er komplekser av tre-fire proteiner utenom rekkevidden til ikke-styrte neo-darwinistiske mutasjoner. De går tapt i form-rommet. Og den store majoriteten av proteiner i celler samarbeider i komplekse sammenkoblinger på seks proteiner eller flere. Langt utenom rekkevidden til ikke-styrte neo-darwinistiske mutasjoner.
Som vi var inne på i punkt 2 (starten): Skal mutasjoner tas vare på -før de har noen gagnlig effekt, skjer ikke det uten en designet mekanisme! Det er hva som kreves for at seleksjons-mekanismen skal virke kumulativt (5). Som evolusjonsbiologen Sir Ronald Fisher har påvist, er det ikke slik i naturen at fordelaktige mutasjoner alltid bevares i en populasjon. De fleste gagnlige mutasjoner blir utryddet av tilfeldige påvirkninger, eller et meget større antall skadelige mutasjoner. Dett er i motstrid med den idé en har hatt siden Darwin at den naturlige seleksjon bevarer den minste gagnlige variasjon inntil den har overtatt hele populasjonen.
Bilde 3. Chlrorquinine-resistens
Klorkinineksempelet viser at det er mulig, men bare i kjempepopulasjoner som det aldri har funnest make til i jordas historie. Alternativet til samtidige mutasjoner, mått f.eks. være det som kalles horisontal genoverføring: NDT postulerer at planter og dyr kan overføre DNA-fragmenter mellom arter, en prosess som kalles 'Horisontal Gen-Overføring' (HGO). HGO er kjent å trekke inn mellom prokaryote celler, slik som bakterier. Bakterielle celler har ingen kjerne til å holde sitt DNA atskilt fra resten av cellen. De kan slynge ut DNA-fragmenter fra en celle til en annen i en form for seksuell overføring eller ved at en bakterie ganske enkelt utskiller et stykke DNA, i form av en genetisk struktur (plasmide), inn i et eksternt medium, som kan bli plukket opp av en annen bakterie. Mye bakterie-resistens vunnet ved patogene bakterier, kommer fra HGO gjennom resistente bakterier.
 Bilde 8. Skisse av prokaryot celle (med kjerne)
Bilde 8. Skisse av prokaryot celle (med kjerne)
Det er imidlertid ingen direkte bevis av slike overføringer mellom ulike arter av eukaryote celler (med kjerne). Jan O. Andersson ved Uppsala Universitet har pekt på at det vi kjenner om gen-overføring eukaryote celler, er basert på uformelle/anekdotiske bevis (Andersson, 2009) Videre i følge Keeling og Palmer (2008), så er eks. med HGO vanligvis identifisert via ulikheter i slektskapstreet mellom arter, ut fra anatomiske og genetiske data. Dermed blir det å forklare ulikheter i fylogenetiske trær ut fra HGO et sirkulært argument. Syvanen, (2012) har uttrykt, uten referanse for øvrig, at HGO er blitt observert i laboratoriet, men ikke så langt i naturlige omgivelser. Dermed har vi ingen direkte bevis på HGO i celler med kjerne. Det er kanskje derfor darwinister foretrekker å forklare problemene ved 'slektskapstreet' ut fra 'konvergent evolusjon'. Om HGO noensinne skulle vise seg å være en viktig faktor i forbindelse med slektskapstrær, så ville hele affæren bli kastet ut i kaos (Syvanen, 2012). I ethvert tilfelle støtter ikke status ved fylogenetiske (slektskaps) -trær oppunder doktrinen om felles avstamning.
Når det gjelder andre mulige årsaker til slike endringer, foreslo Dr. Spetner En Ikke-Tilfeldig Evolusjons-Hypotese, fordi den kunne redegjøre for raske evolusjonære endringer som har funnet sted. Evnen til å gi tilsvar til endringer i omgivelsene, krever en organisme som er i stand til å oppfatte en endring i omgivelsene og har en mekanisme hvorved denne sansede endringen fører til aktivitet av et latent gen, eller andre genetiske ressurser. Disse vil i sin tur lede til endring i fenotypen, som gir organismen en fordel i de nye omgivelsene. B. Sanner ved Emory Universitet har foreslått noe likt dette for bakterier, at genomisk rearrangement kan trigges av endringer i omgivelsene via et kontrollsystem i bakterien som kan gi arvelige endringer (Wanner, 1985). Chr. Cullis og kolleger ved CaseWestern Reserve Universitet i Cleveland har rapportert at miljøendringer framkaller genetisk rearrangement i linplanter. Det er de samme genetiske endringene som inntreffer når de samme miløendringer inntreffer, noe som indikerer at genetiske endringer stimuleres av omgivelsene ikke ved tilfeldigheter. (Cullis 2005, Chen et al. 2009) Men forfatterne vil ikke være med på designede mekanismer i livets maskineri. Det meste må stemme med 'vanlig vitenskap'. Fagvitenskapen bestemmer, men den synes ikke å fungere gjennom å få sine hypoteser og bekreftet i laboratoriet, eks. her. Andre mulige teoretiske forklaringer er selvsagt mulige, men har ikke kjente praktiske utløpere/konsekvenser.
I en simulering utført av Lenski (2003 ) fant man at om ingen selektiv fordel foreligger helt til funksjonalitet er oppnådd, så kan ikke evolusjon produsere det endelige funksjon.
Konseptet med å selektere på individer med flest mutasjoner kan fungere i meget begrenset grad i naturen, men det er klart ikke det som vanligvis skjer. En overveldende variasjon i evne til å fungere (fitness) forårsakes av noen få spesifikke og høyst kraftige genetiske variasjoner, og ikke av det totale mutasjonsantallet pr. person. Som eks. kan nevnes at 3 koordinerte mutasjoner vil kreve flere milliarder år, om det skal skje tilfeldig. Det kan empiri f.eks. i forbindelse med malaria, og resistens mot denne
Som svar på 'det store spørsmålet': 'Hva eller hvem skylder vi vår eksistens?' -så er det knapt noe menneske som våkner opp om morgenen og takker naturlig seleksjon eller blinde repetisjoner for sin eksistens. Men om vi ikke er avkom fra noen utilsiktet årsak i det hele, så blir derimot hensikt en nøkkel-ingrediens i vårt opphav.
Referanser:
1. O. Khersonsky og D.S. Tawfik, "Enzyme Promiscuity: A mechanistic and Evolutionary Perspective," Annual Review of Biochemistry 79 (2010): 471-505
2. R. Mukhopadhyay, "Close to a Miracle: Researchers are debating the Origins of Proteins, ASBMB Today 12, no. 9 (2013): 12-13
3. A.K.Gauger og D. Axe, "The Evolutionary Accessibility of New Enzyme Functions: A Case Study from the Biotin Pathway," BIO-complexity, no I (2011): 1-17
4. M.A. Reeves, A.K. Gauger og D. Axe, "Enzyme Families: Shared Evolutionary History of Shared Design? A study of the GABA-Aminotrarnsferase Family," BIO-Complexity, no. 4 (2014), 1-16
5. Life's irreducible Structure. Science 160, 1968, s.1309
Selektert og samlet av Asbjørn E. Lund
{kind=link}
{kind=link}